近日,中国科学院南海海洋研究所喻子牛研究员团队鹦鹉螺基因组研究成果以“The genome ofNautilus pompiliusilluminates eye evolution and biomineralization”为题,在国际著名期刊Nature Ecology & Evolution(IF:12.544)上在线发表,百迈客生物科技作为共同一作共同参与研究。文中通过三代PacBio技术成功构建了完整的鹦鹉螺基因组,并揭示了鹦鹉螺针孔眼形成和生物矿化的潜在机制。本研究为现存的头足类动物重建进化情景和基因组变化特征提供了宝贵资源。


研究背景
鹦鹉螺是古生代以来灭绝的数百个头足类属中唯一幸存的外壳头足类,其在漫长的演化历程中依然保留了祖先特征,如螺旋外壳和针孔眼等。鹦鹉螺壳室为黄金比例的对数螺旋形式,由坚固的文石晶体阵列组成,导致其高度的静水稳定性;鹦鹉螺独特而简单的针孔眼,没有晶状体或角膜,为阐明眼睛的进化提供了极好的原型模型。
由于观赏性的商业开发、缺乏法律保护及性成熟缓慢等诸多不利因素,鹦鹉螺种群数量急剧下降。因此,研究鹦鹉螺的基因组可以为头足类动物的进化驱动因素提供见解,并为物种保护提供重要理论基础。

主要研究结果
1.鹦鹉螺基因组构建
通过81.8X Illumina + 112.5X PacBio,组装730.58 Mb鹦鹉螺基因组(survey预估753.09 Mb),contigN50=1.1Mb,BUSCO评估为91.31%。鹦鹉螺是迄今为止被测序的头足类动物中基因组最小的物种 (图1)。通过多种方式注释17,710个编码蛋白基因(BUSCO为93.46%),基因数为章鱼和鱿鱼的52.6–60.5%。
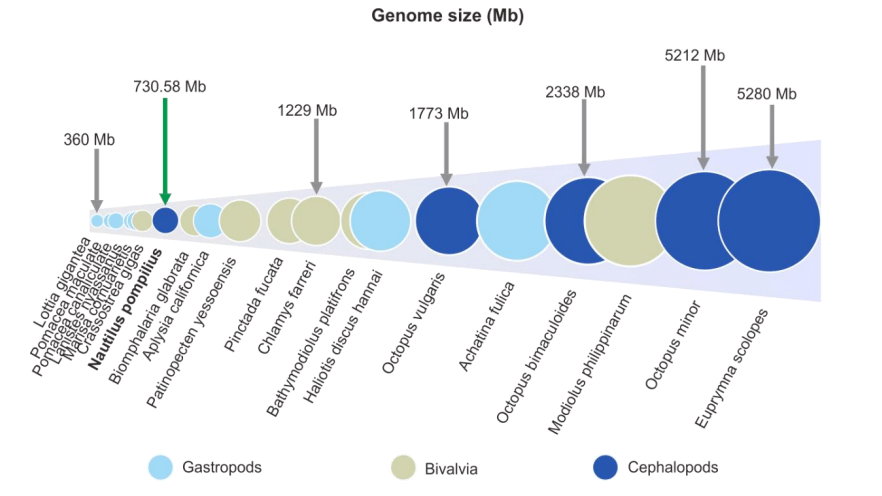
图1软体动物基因组大小分布图
包括转座子(transposable elements,TEs)在内的重复因子是构成基因组结构和进化的驱动力,比较分析进一步发现鹦鹉螺中TEs的组成与coleoid谱系明显不同(图2a)。在头足类动物中可以观察到逆转录转座子(LINE和LTR)爆发,鹦鹉螺基因组中没有最近的TEs扩增(图2b),证实了反转录转座子在推动其基因组进化中的关键作用。并 通过Tajima’s相对速率检验等方式检测发现鹦鹉螺编码区的进化速率较慢。
基因家族分析显示鹦鹉螺中发生了大量的收缩(204个基因家族收缩,9个基因家族扩张)。对神经系统的神经发生和适应性具有功能意义的锌指转录因子和原钙粘蛋在章鱼基因组中大量扩张,而鹦鹉螺基因组中没有。鹦鹉螺基因组中特异性扩增了18个着丝粒蛋白B(CENPB)结构域的基因。CENPB通过抑制酵母或人类的反转录转座子和着丝粒的形成,在宿主基因组的完整性和进化中起着重要的作用。因此,CENPB扩增可能是维持古基因组完整性的因素。
2.系统发育分析和种群规模估计
对16个头足类物种基因组构建进化树,结果证实鹦鹉螺是coleoids的姐妹类群,它们的分化时间在志留纪-泥盆纪边界附近(4.226亿年前)(图2c),古软体动物的分化和物种形成始于埃迪卡拉纪时期,在此期间,早期后生动物出现了逐渐多样化和生物新颖性。
种群历史动态分析显示,在过去的几百万年中,两个重要的环境演化事件对鹦鹉螺 种群的形成产生了深远的影响。特别是在中新世之交(22.6百万年前),鹦鹉螺种群呈逐步扩大的趋势。然而它们的上升在中更新世过渡的早期阶段停止,这与基本的气候变化相一致。
在38万年前鹦鹉螺种群数目锐减(接近于40万年前左右的中布容事件(MBE)),表明鹦鹉螺对海洋环境变动非常敏感。同时,MBE也是夏威夷短尾乌贼( Euprymna scolopes)和真蛸(Octopus vulgaris)等coleoid动物种群扩张的一个转折点(图2d)。
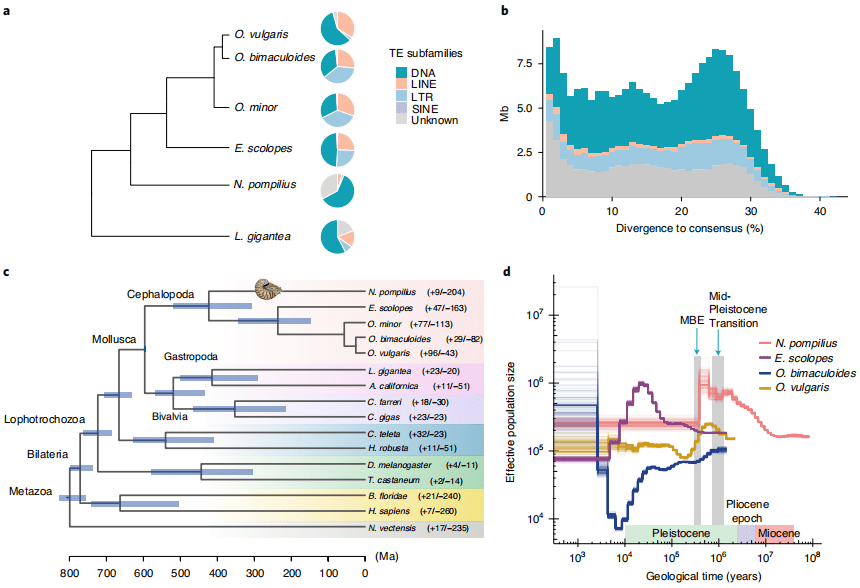
图2 鹦鹉螺基因组结构及与头足类进化
3.Homeobox基因聚集分析
Coleoids的一个突出改变是失去外壳,外壳被内化为一个浮力补偿装置,推动了其显著的多样化。同源异型盒(Homeobox)基因是后生动物进化过程中躯体发育和组织划分所必需的关键转录因子,因此通过比较多个谱系中Hox簇的组织结构,探讨头足类体型进化的遗传基础具有重要的意义。研究结果显示鹦鹉螺基因组包含一套完整的软体动物Hox基因(图3)。而夏威夷短尾乌贼中的Hox2缺失、双斑蛸(Octopus bimaculoides)中的Hox2–Hox4缺失、加利福尼亚海兔(Aplysia californica;一种没有外壳的腹足动物)Hox2,Hox4和Antp缺失(图3),表明Hox簇完整性的破坏可能与软体动物谱系中外壳的进化丢失有关。
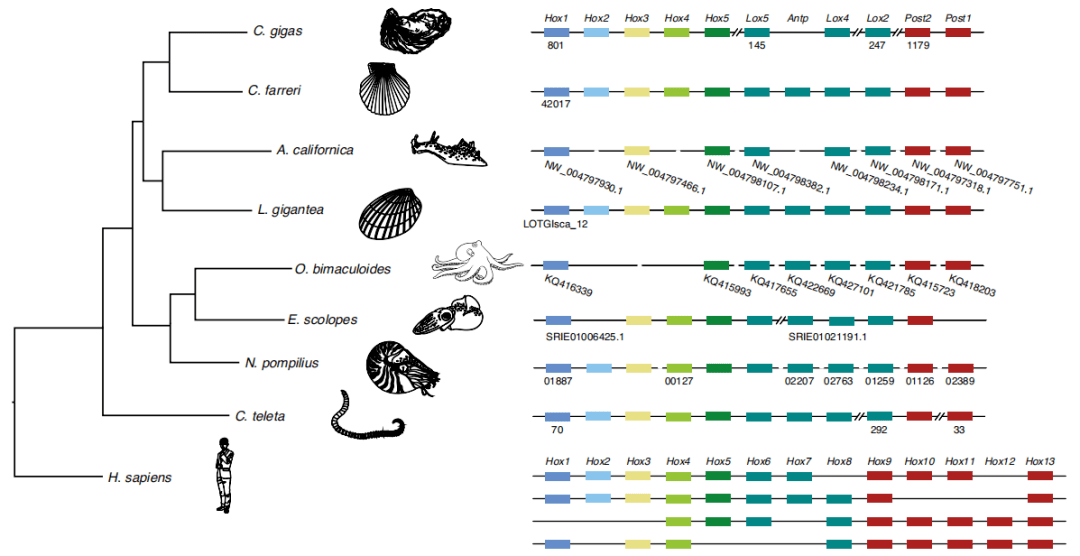
图3 后生动物基因组中Hox基因簇的示意图
4.针孔眼的演变
针孔眼是鹦鹉螺最奇特、最显著的特征之一,在鹦鹉螺的视网膜上,用可调节的瞳孔代替晶状体形成一个相对暗淡的图像。与复杂的Coleoid照相眼睛相比,针孔眼相对简单的结构是讲述眼睛和/或晶状体形成起源的良好模型。对控制晶状体形成的核心调控转录因子的基因组搜索显示,几乎所有这些核心调控因子,包括PAX6、SIX3/6和SOX2,都存在于鹦鹉螺基因组中(图4a),与此前古生物学研究报道相符(早寒武世就出现了带有晶状体的眼睛化石),结果支持了晶状体的古老起源。
系统发育分析表明,软体动物Nrl/Mafa-Mafc属于大型Maf超家族,其直系同源物在脊椎动物中可分为Mafa、Mafb、c-Maf和Nrl四个分支(图4b)。 Maf大家族成员在表达上具有晶状体特异性的观点,并且在脊椎动物的晶状体诱导和分化中起着核心作用。Nrl或c-Maf的补充可以增强PAX6诱导的晶体蛋白(光折射所需的最丰富的晶状体结构蛋白)。比较结果表明,鹦鹉螺基因组中的 Nrl/Maf基因具有谱系特异性缺失(图4b)。在coleoids中,S-晶体蛋白谱系特异性扩增,而在鹦鹉螺基因组中没有S-晶体蛋白基因(图4c)。对晶体蛋白近端上游序列转录调控位点的研究表明,coleoids中的NRL/MAF结合motif比在鹦鹉螺中更为丰富。以上结果说明,鹦鹉螺基因组中晶状体发育的关键转录因子NRL/Maf的缺失和晶状体形成蛋白的基因家族收缩,共同促进了鹦鹉螺针孔眼的形成。
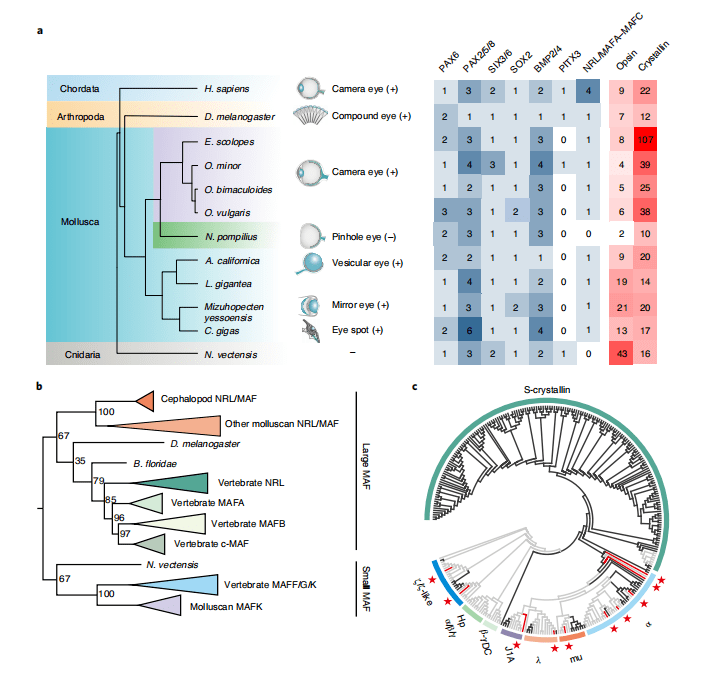
图4 NRL的缺失和晶体基因的收缩与针孔眼的进化相关
鹦鹉螺是一种夜行性食肉动物,具有夜间向浅水垂直深度迁移的特征 ,光感应和空间视觉是完成这项任务的基本前提。系统发育证据表明鹦鹉螺基因组编码一个光受体视紫红质基因和一个视黄色素基因,是已知后生动物中最小的视紫红质基因数目(图4a)。表达模式分析显示r-视蛋白(r-opsin)及其相关信号级联主要在眼睛中表达(图5),表明r-视蛋白(r-opsin)的主要作用在于介导鹦鹉螺 中的横纹肌节光转导。鹦鹉螺仅保留最简单的光信号传导通路和唯一的感光分子r-opsin,使其缺乏对颜色的分辨能力,成为“色盲”。
由于深海水域亮度的急剧下降,对垂直迁移的海洋动物来说,光强度的感知更为关键 。视蛋白对光的敏感性很大程度上取决于11-顺式视网膜的生色团,其异构化通常导致构象变化和视蛋白信号转导的激活。在头足类中,视网膜色素是视觉循环中一种主要的和谱系特异性的异构酶。在脊椎动物中,视网膜色素上皮特异性蛋白65kDa(RPE65)是一种关键的异构酶,通过将全反式视黄酯转化为11-顺式视黄醇来驱动视黄酸循环。在鹦鹉螺基因组中发现并鉴定了RPE65基因家族的扩张,该家族共编码10个基因。鹦鹉螺RPE65与人RPE65共有一个保守的铁离子结合位点、一个活性位和一个用于进入基质的疏水通道,说明是具有潜在的催化活性。从进化适应的角度来看,针孔眼的出现是鹦鹉螺垂直深度迁移生活方式的一个适应性突破,通过将瞳孔打开,让生物体获得空间视觉并快速应对眼睛内的静水压力。总的来说,包括基因缺失、特定基因家族的独立收缩和扩张以及相关调控网络的存在在内的多种基因组共同推动了鹦鹉螺针孔眼的进化。
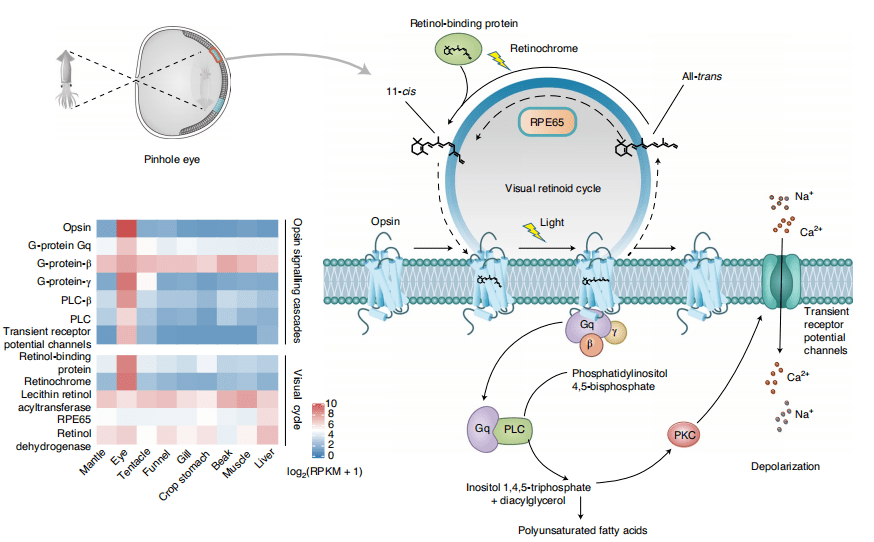
图5 鹦鹉螺的视觉模型
5.壳的形成
鹦鹉螺是现存唯一具有外骨骼的头足类动物,它拥有复杂的螺旋形外壳,不仅是抵御捕食或环境逆境的物理屏障,而且在维持浮力方面起着不可或缺的作用。因此,鹦鹉螺独特的贝壳结构是垂直迁移适应性进化的结果。一般来说,软体动物贝壳的形成是一个基本的生物矿化过程,其中贝壳基质蛋白(SMPs)引导碳酸钙多晶型(方解石和/或文石)的生长,并将晶体组织成复杂的贝壳。了解鹦鹉螺壳层的超微结构结构和SMP生物组成对于揭示壳层形成和演化的古老机制非常重要。作者通过扫描电子显微镜(SEM)图像揭示了沿生长方向堆积的六方文石的纯聚集体(图6a)。文石可能是在方解石成为构建软体动物外壳的主要材料之前的古代结晶碳酸钙。为了进一步研究鹦鹉螺贝壳形成的分子基础,从2个技术重复的酸溶性(ASM)或酸不溶性(AIM)基质组分中鉴定出78种SMPs(图6b)。表达模式显示,这些SMPs中的大多数(72.2%)在套膜中表达特别高,从而证实了套膜在贝壳形成中的中心作用。为了描述保守的软体动物生物矿化“toolkit”,作者进行了比较壳蛋白质组学分析,结果表明,鹦鹉螺21种SMPs与其他软体动物(包括双壳类和腹足类)的对应物具有相似性(图6c)。进一步的结构域分析揭示了软体动物的几个保守结构域,包括Sushi/SCR/CCP、层粘连蛋白、几丁质结合和碳酸酐酶结构域。表明这些结构域可能作为一种古老的“核心生物矿化工具包”出现,并在具有外壳的多种软体动物谱系中保存。OrthoFinder分析显示,78种SMPs中有52种提供了新的或鹦鹉螺特异性的壳蛋白(图6d),从而推测大多数独特的SMPs是独立进化的,并有助于软体动物壳结构的高度多样性。还发现鹦鹉螺壳基质蛋白存在全新的低复杂重复基序(RLCDs)(图6e),这可能与鹦鹉螺独特的壳结构相关。另外,系统比较软体动物壳基质蛋白发现,RLCDs的平行进化可能是软体动物生物矿化的基本规律。
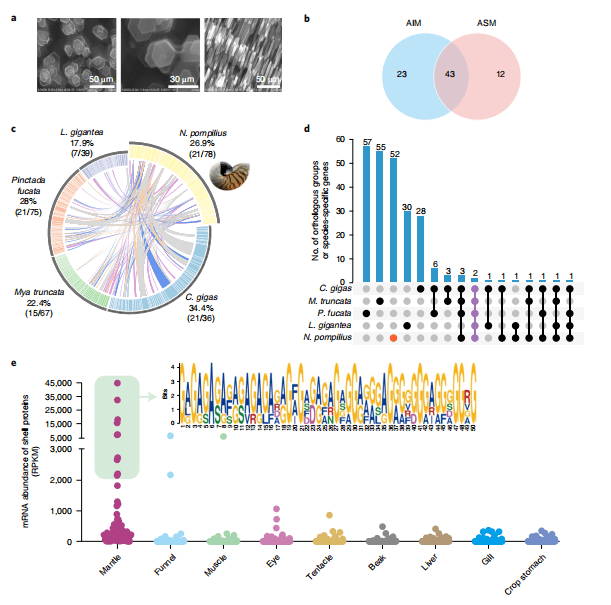
图6 鹦鹉螺壳的超微结构和蛋白质组
6.免疫系统
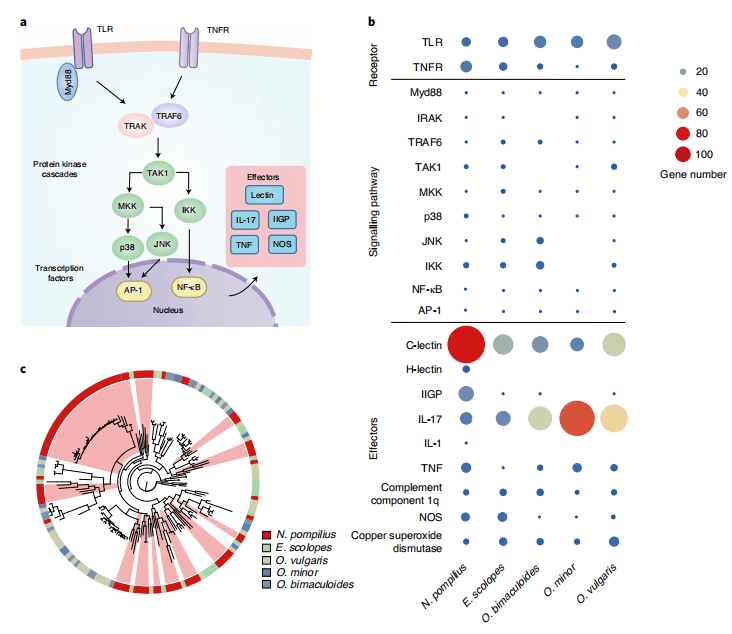
图7 鹦鹉螺免疫系统中功能完整和特异的基因扩增
小结
本研究通过鹦鹉螺基因组测序和进化分析,揭示了针孔眼形成和生物矿化的演变规律,并结合转录组、蛋白组、基因家族及系统发育等分析对针孔眼、壳形成机制进行了深入剖析。相关的研究成果对鹦鹉螺的生物学研究、遗传资源保护以及头足动物的适应性演化都将发挥重要的推动作用。
文:豆沙包
排版:市场部
百迈客生物基于高通量测序技术、生物信息分析技术和生物云计算技术,为广大科研工作者提供以综合技术服务、生物云分析、三代高通量测序以及试剂、仪器等科研周边业务。
转自:百迈克生物- 本文固定链接: https://maimengkong.com/zixun/1448.html
- 转载请注明: : 萌小白 2023年4月16日 于 卖萌控的博客 发表
- 百度已收录
