来自美国加州大学的冯根生团队,利用scRNA-seq绘制了从新生儿到成年的单细胞分辨率小鼠肝脏发育图谱,证明了肝细胞与内皮细胞、星状细胞和Kupffer细胞协同作用的功能成熟,以及巨噬细胞群的短暂出现。这项研究提供了一个全面的图谱,涵盖了所有的肝细胞类型,并有助于进一步剖析肝脏发育、代谢和疾病。该文章在2022年2月7日发表在Developmental Cell,以下是文章的详细解读。

文章题目:Temporal Analyses of Postnatal Liver Development and Maturation by Single-Cell Tranomics
发表时间:2022-02-07
发表期刊:Developmental Cell
主要研究团队:美国加州大学等
影响因子:13.417
DOI:10.1016/j.devcel.2022.01.004
研究背景
肝脏执行重要的功能,包括新陈代谢、解毒、胆汁分泌和血浆蛋白的产生。肝细胞是主要的实质细胞,占肝脏体积的80%,并执行大部分的代谢和合成功能。在成人肝脏中,最基本的构筑单位是条索状肝小叶,其中门静脉(portal vein,PV)、肝动脉和胆管在六边形小叶的拐角处形成一个门静脉三角,中央静脉(portal vein,CV)位于小叶中心。血液从PV和肝动脉进入小叶,在流经血窦时与肝细胞交换营养物质和氧气。肝细胞异质性伴随着氧气、营养物质和激素的梯度,在CV和PV区域之间形成代谢区。位于小叶轴不同层次的肝细胞表达不同的受体、转运蛋白和酶。出生后,肝细胞组织成肝小叶,形成分带结构。然而,带状结构形成的驱动因素,以及肝细胞如何采取特定的代谢功能仍有待确定。大数据分析只能提供不同细胞类型的平均值,而混淆了细胞类型组成变化、细胞-细胞相互作用和功能异质性的关键信息。综合利用scRNA-seq和单分子荧光原位杂交,允许在禁食的成年小鼠中重建肝脏分区和推断小叶坐标,超过一半的肝脏基因显示显著的分区,尽管分析集中在肝细胞。
研究样本
scRNA-seq样本:出生后第1、3、7天(D1、D3、D7)的新生样本,每个时间点包括4只雄性小鼠;出生后第21、56天(D21、D56)的样本,每个时间点纳入2只雄性小鼠。
FISH:D7和D21小鼠新鲜肝脏冷冻切片。
研究策略
研究人员首先分离了所有主要的肝细胞类型,以单细胞分辨率对小鼠出生后的肝脏发育进行全面分析。然后,对肝脏中昼夜节律的发展,以及出生后肝细胞中几种关键代谢途径的反应进行了计算预测,包括糖酵解、脂肪酸氧化、脂质和胆固醇的生物合成。最后,研究人员鉴定了一组在出生后第7天富集的特殊的巨噬细胞,并通过细胞-细胞通信分析推断了它们在调节窦状血管化和调节T(Treg)细胞活性中的作用。
研究成果
1. scRNA-seq在出生后早期肝脏中识别出不同的肝细胞亚群
收集了小鼠出生后D1、D3、D7、D21和D56的细胞,这些细胞覆盖了从新生到成年的整个肝脏发育时期。所有分离的单细胞进行scRNA-seq分析,共获得52,834个转录组。研究人员使用UMAP对来自5个时间点的细胞进行归一化处理,并进行了聚类和降维分析(图1A、1C)。根据人工挑选的已知标记的表达情况,从所有年龄中共鉴定出31种细胞类型(图1B)。
研究人员对来自D1到D56的9,137个肝细胞进行研究,它们表达生物标记物Alb和Hnf4a (图1B)。这些细胞主要按时间点聚集(图1D),相对于D21和D56细胞,来自D1、D3和D7的肝细胞更靠近。一些肝母细胞或肝细胞癌(hepatocellular carcinoma,HCC)标志物,如Afp、Ahsg和H19,在早期时间点被肝细胞高表达(图1E),表明它们不是成熟的肝细胞。事实上,D1~D7的肝细胞正在积极增殖,高表达Mki67,编码增殖标志物Ki67(图1F)。来自D21的肝细胞处于过渡状态,Mki67、Afp、Ahsg和H19低表达(图1E和1F),Hnf4a、Cyp1a2和Cyp3a11也低表达(图1G),成熟肝细胞标志物在成年肝脏中表达(D56)。
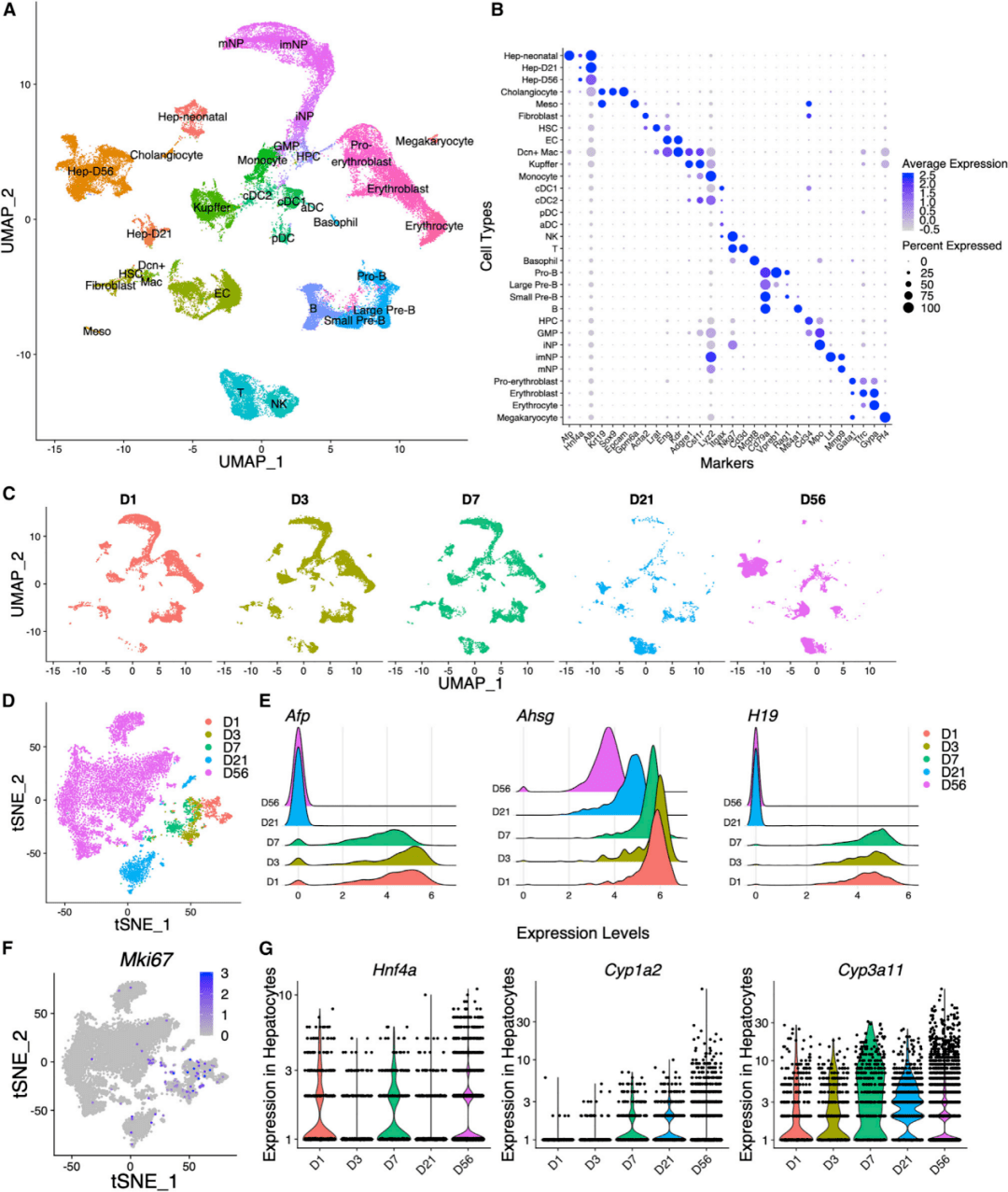
图1 发育中和成年小鼠肝脏中的肝细胞类型
在每个时间点,研究人员鉴定了2~3个亚群(图2A~2E),其中有几个标记基因在多个时间点共有。D1-Hep2、D3-Hep2和D7-Hep2高表达代谢相关基因Cyp4a14和Cps1(图2A~2C)。在后期,D21-Hep3和D-56Hep3表达Snca和Cd24a(图2D、2E)。有趣的是,D1-Hep3具有Scd2的独特表达特征(图2F),它编码硬脂酰CoA去饱和酶的一个异构体——单不饱和脂肪酸生物合成的关键酶。Scd1在D21和D56表达量较高,而Scd2主要在D1表达(图2G、2H)。研究人员通过qRT-PCR在分离的肝细胞中验证了Scd1和Scd2不同的表达模式(图2I、2J)。
在D21和D56中鉴定出2个罕见的肝细胞亚群(图2D、2E)。研究人员观察到,在Hep1和Hep3细胞中,Malat1表达与mtRNA比例高度相关,而在Hep2中则不相关(图2K) ,支持Hep2不是一群低质量细胞的观点。接下来,研究人员在D7和D21对Neat1和Malat1进行小分子荧光原位杂交(smFISH)(图2L)。Pearson's相关分析显示,Malat1和Neat1的表达高度相关,在D21确定一组肝细胞以它们的共表达为特征。
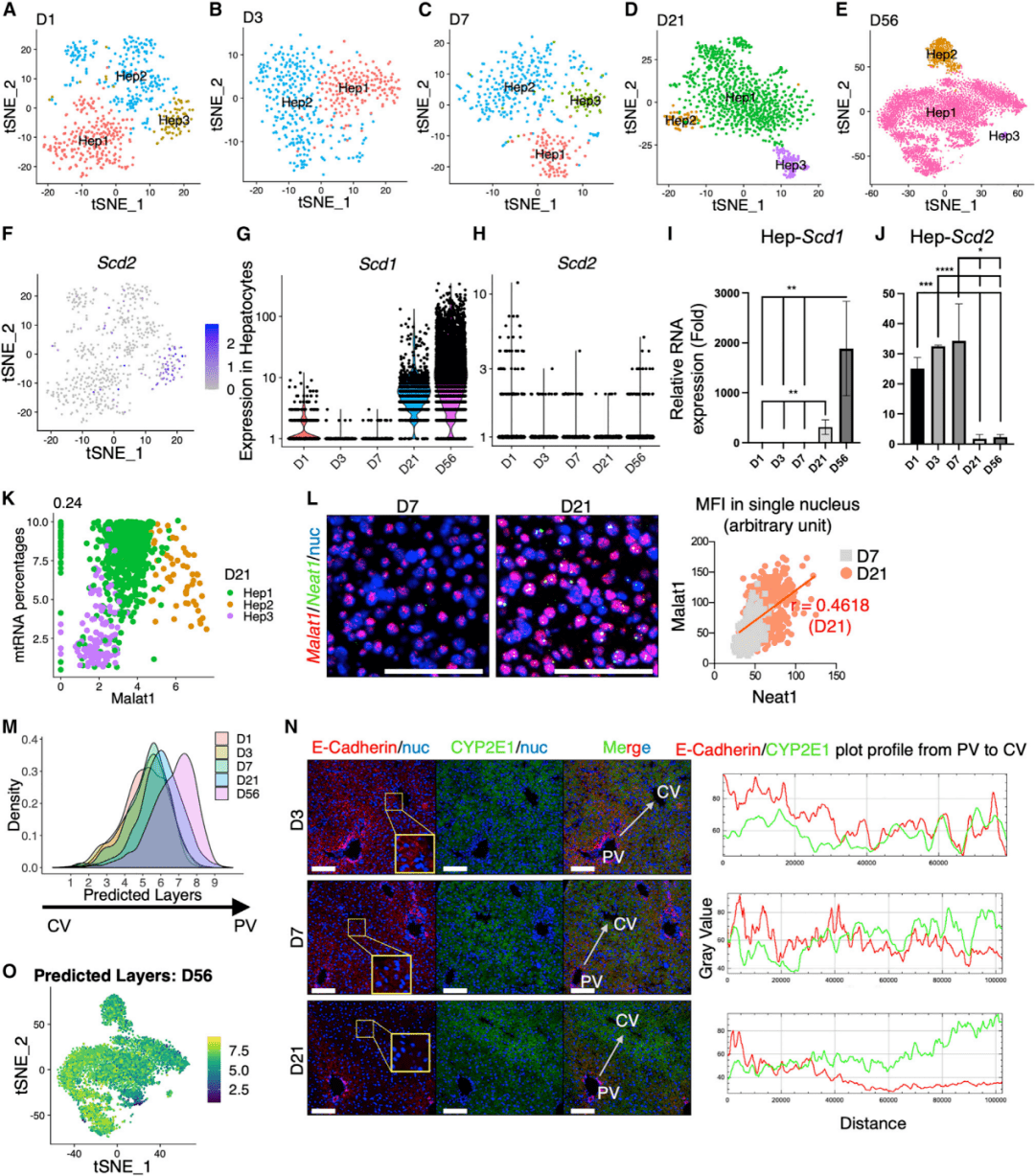
图2 每个时间点肝细胞中不同的转录组图谱
2. 逐步建立沿小叶轴的分区模式
先前的一项研究根据scRNA-seq和smFISH数据将肝小叶分为9层。在此,研究人员用指定的层训练该数据集,并从肝细胞簇中推断出单个细胞的空间位置。第1层代表CV区的肝细胞,第9层接近PV。预测的D56的层分布(图2M)与所提出的成人肝脏分区模型的先验概率一致,密度在第8层达到峰值。然而,在早期时间点预测的分区没有显示出类似的模式(图2M),表明出生后肝脏中渐进的分区构建。肝细胞中18个最显著的分区标志基因的热图显示,从D1到D56,CV到PV逐渐转变。E-cadherin和CYP2E1的免疫染色始终显示,PV和CV肝细胞的表达相互增加。研究人员将预测的层映射回了D21和D56的tSNE可视化(图2O)。虽然与D3和D7相比,免疫染色在D21的CV和PV区域中检测到E-cadherin和CYP2E1有相反的表达模式(图2N),转录组分析显示,大多数来自不同层的肝细胞仍混合在D21,提示D21肝细胞的代谢区域尚未完全发育。与区域标记物的表达一致,分区肝细胞只能在D56被区分(图2O)。
研究人员使用Monocle建立了发育轨迹,并计算了每个单细胞的拟时序(图3A、3B)。将D56肝细胞分为三组:D56-p.c.、D56-mid、D56p.p.,根据预测层和随机降采样,与其他时间点进行比较。由于从D1到D56收集了肝脏样本,实时点可以用来评估拟时间计算的准确性(图3C)。在D56,近中心的肝细胞(D56-p.c.)首先出现在运动轨迹上,其次是小叶中部(D56-mid),再次是汇管区(D56-p.p.)。与之一致的是,分区预测也显示门静脉周围肝细胞向D56方向稳定增长(图2M),表明门静脉周围肝细胞功能成熟较晚。
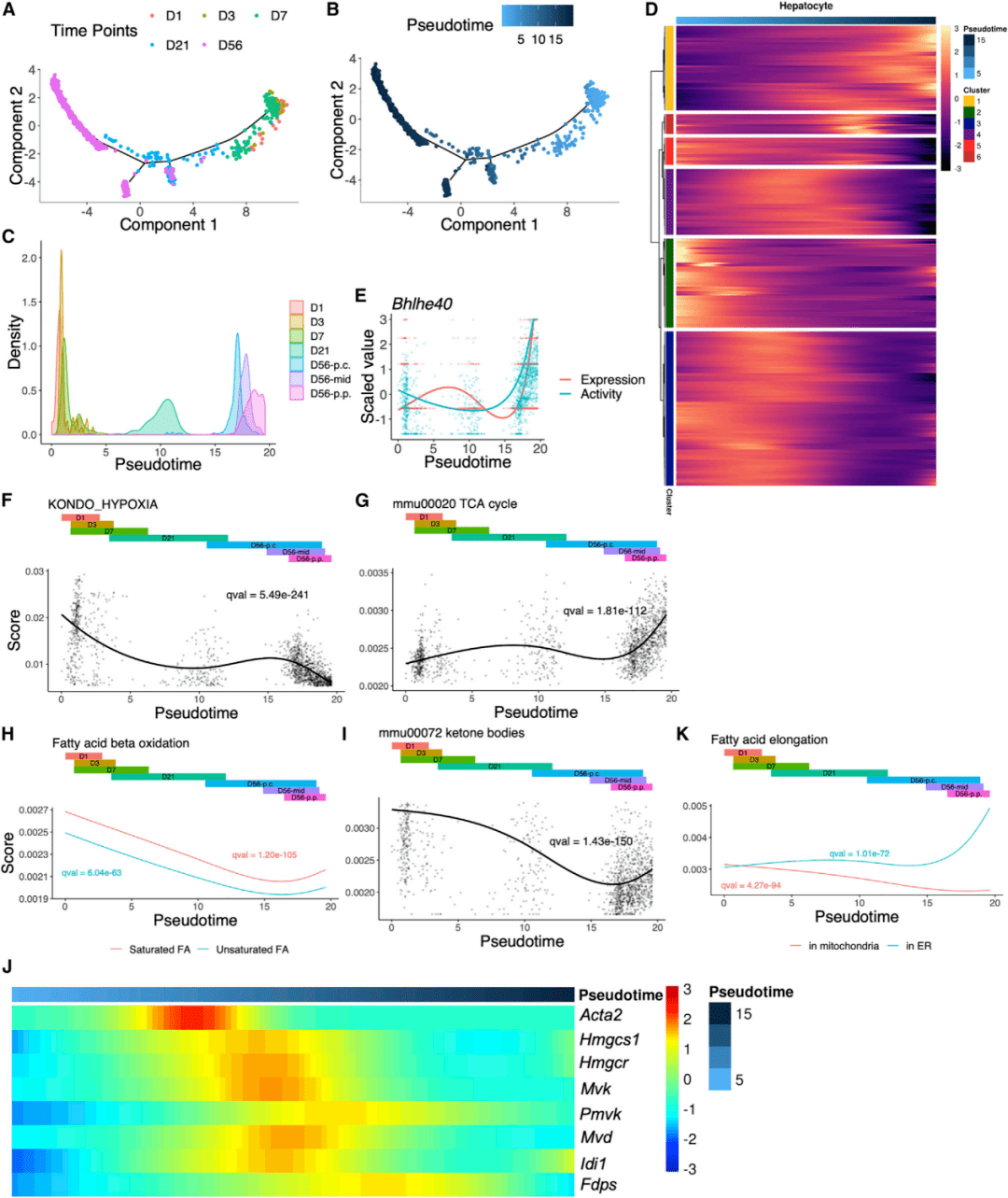
图3 肝细胞内转录因子活性和代谢功能的动态变化
3. 肝脏发育过程中转录因子的鉴定及代谢功能分析
研究人员通过拟时间对所有时间点的单个肝细胞进行了排序,并进行差异表达分析,以确定表达显著变化的基因作为拟时间的功能。然后,这些选定的基因被分组为6个簇的伪时间表达模式(图3D)。在cluster 1中发现了外周生物钟相关通路,其中基因从D1到D56逐渐增加。通过单细胞调控网络推断和聚类,研究人员捕获了36个转录因子(TFs), 它们的表达和活性随着拟时间的变化而发生显著变化,包括昼夜节律的核心调控因子Bhlhe40(Dec1)。其表达和活性在后期急剧增加(图3E),与通路富集数据一致(图3D)。
为了探索肝细胞中代谢功能的发育,研究人员计算了从KEGG(the Kyoto Encyclopedia of Genes and Genomes)数据库和MSigDB收集的基因集的通路富集分数,然后沿着拟时间对细胞进行排序,并筛选出得分沿轨迹显著变化的基因集。研究人员发现缺氧相关通路的表达从D1到D56持续下降(图3F)。在D56时,缺氧通路在PV中比在CV区域更明显地被抑制,而在三羧酸循环中观察到相反的模式(图3G)。而脂肪酸β-氧化在出生后立即出现一个高峰,随后随着时间的推移迅速下降,并在D56观察到门静脉周围肝细胞略有增加(图3H),产生大量的乙酰辅酶A。酮体相关通路表现出与β-氧化类似的模式,在出生后立即达到峰值(图3I)。参与胆固醇生物合成的甲羟戊酸途径的基因在新生儿中适度表达,但在D21达到峰值(图3J),表明该通路在新生儿肝脏中建立不完善。线粒体中的短脂肪酸延伸可能对新生肝脏中的肝细胞很重要,尽管内质网在发育后期主导了这一过程(图3K)。
4. 肝内皮和间充质细胞的发育
所有五个时间点的肝脏内皮细胞(endothelial cells,ECs)都聚集成11个亚群(图4A),随着时间的推移,非肝窦内皮细胞(liver sinusoid endothelial cells,LSECs)共享类似的转录组。D56的LSECs构成2个大而连续的群体,分别为近汇管区和近中心的LSECs(EC1和EC2)(图4B)。相反,研究人员在未成熟LSECs(EC3~EC6)中观察到更高的异质性,没有明显的分区分离。在所有时间点检测到一小群表达增殖和细胞周期相关基因的EC9细胞,但呈减少的趋势(图4B)。根据细胞周期标志基因预测,所有EC9细胞均处于G2/M或S期,表明在发育中的肝脏中存在一群具有增殖潜能的LSEC。
通过将胚胎与本研究中新生儿内皮细胞结合数据(图4C、4D),研究人员构建了一条从E7.5到D7的轨迹(图4E)。分析表明,PV和CV区的内皮细胞采取不同的发育路径,PV区在出生后被重塑,该区域的肝细胞和内皮细胞协同分化。采用与肝细胞类似的方法,研究人员构建了LSECs的发育轨迹,并计算了每个单细胞的拟时序(图4F)。这种方法确定了包括Bhlhe40、Tcf4、Hmgb1和Meis2在内的多个转录因子在LSEC发育过程中表现出明显的表达和活性变化(图4G)。接下来,研究人员通过分离造血干细胞、成纤维细胞和间皮细胞,鉴定了更多的间充质细胞亚群(图4H),这些细胞亚群最初包含在其他间充质细胞中。Meso1和Meso2两个亚组尽管具有不同的转录组图谱,但有共同的间皮标志物Gpm6a、Upk3b和Wt1(图4H)。Alcam是小鼠肝脏中胚胎间皮细胞的标志物,在Meso1中表达,但在Meso2中不表达。值得注意的是,Meso1主要由新生期的细胞组成(图4I和图4J)。
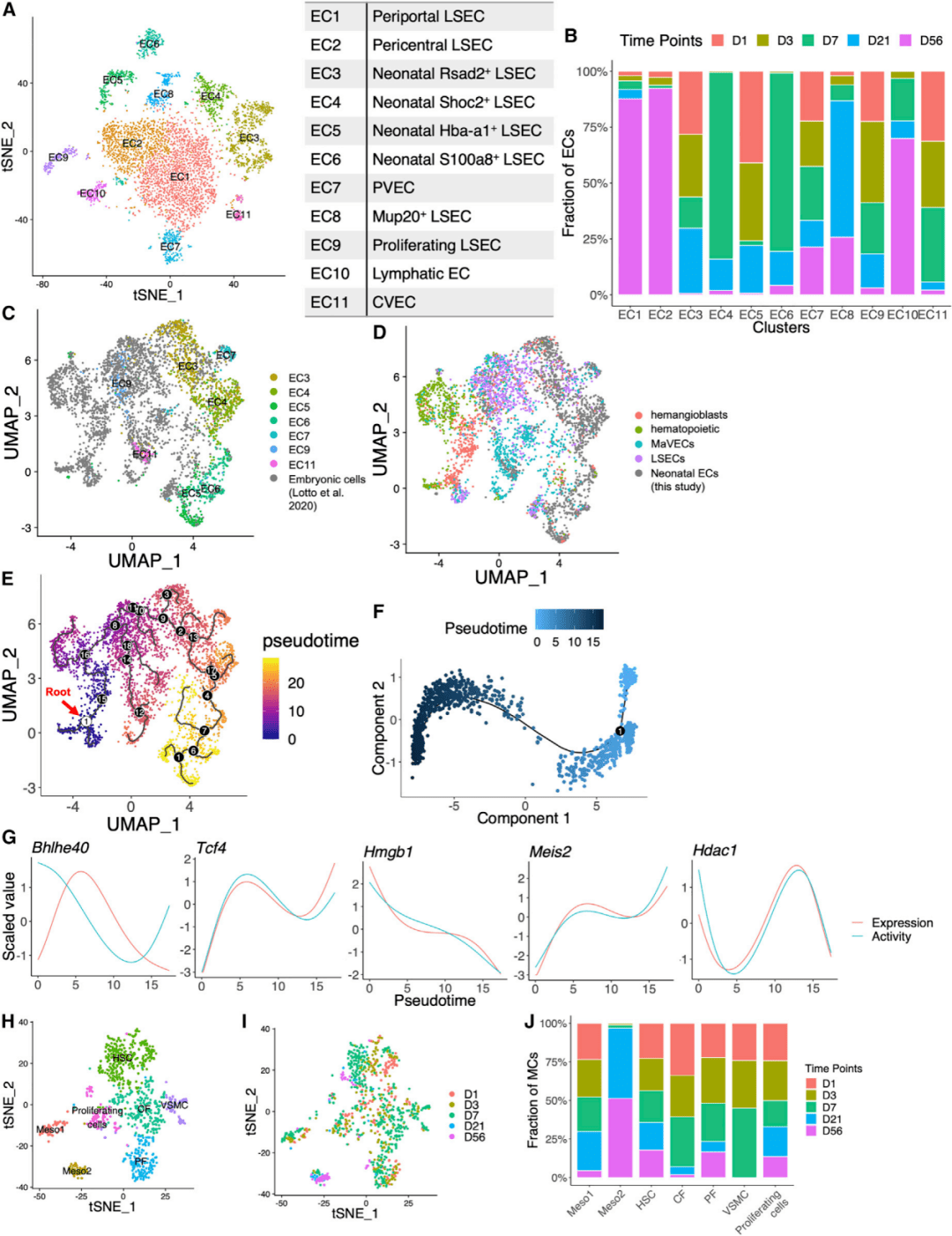
图4 肝脏内皮和间质细胞的发育
5. 造血和免疫细胞群的动态变化
从D1到D56的数据集显示了许多发育中的造血细胞,包括造血祖细胞(hematopoietic progenitor cell,HPCs)、粒细胞-单核细胞祖细胞(granulocyte-monocyte progenitors,GMPs)、B细胞、中性粒细胞和红细胞谱系(图1A)。HPCs和GMPs均表达造血前体细胞标志物Cd34(图1B)。通过力导向图分析推断了这些细胞类型的发育轨迹(图5A),证实了UMAP可视化显示的预测关系(图1A)。HPCs连接3条主要通路,分别向GMPs、红细胞谱系和B细胞方向发展。随后,GMPs分化为单核细胞和中性粒细胞。肝造血功能在出生后持续存在,但迅速消退,在D7后几乎检测不到(图5B)。在鉴定的11种T和NK细胞亚型(图5C)中,Treg细胞(图5D)显示出随时间的独特变化,在D7达到峰值(图5E)。研究人员比较了D7、D21和D56之间在Treg细胞中差异表达的基因,发现排名前两位的基因是Brd1和Gtf2h3(图5F),可能参与细胞增殖。
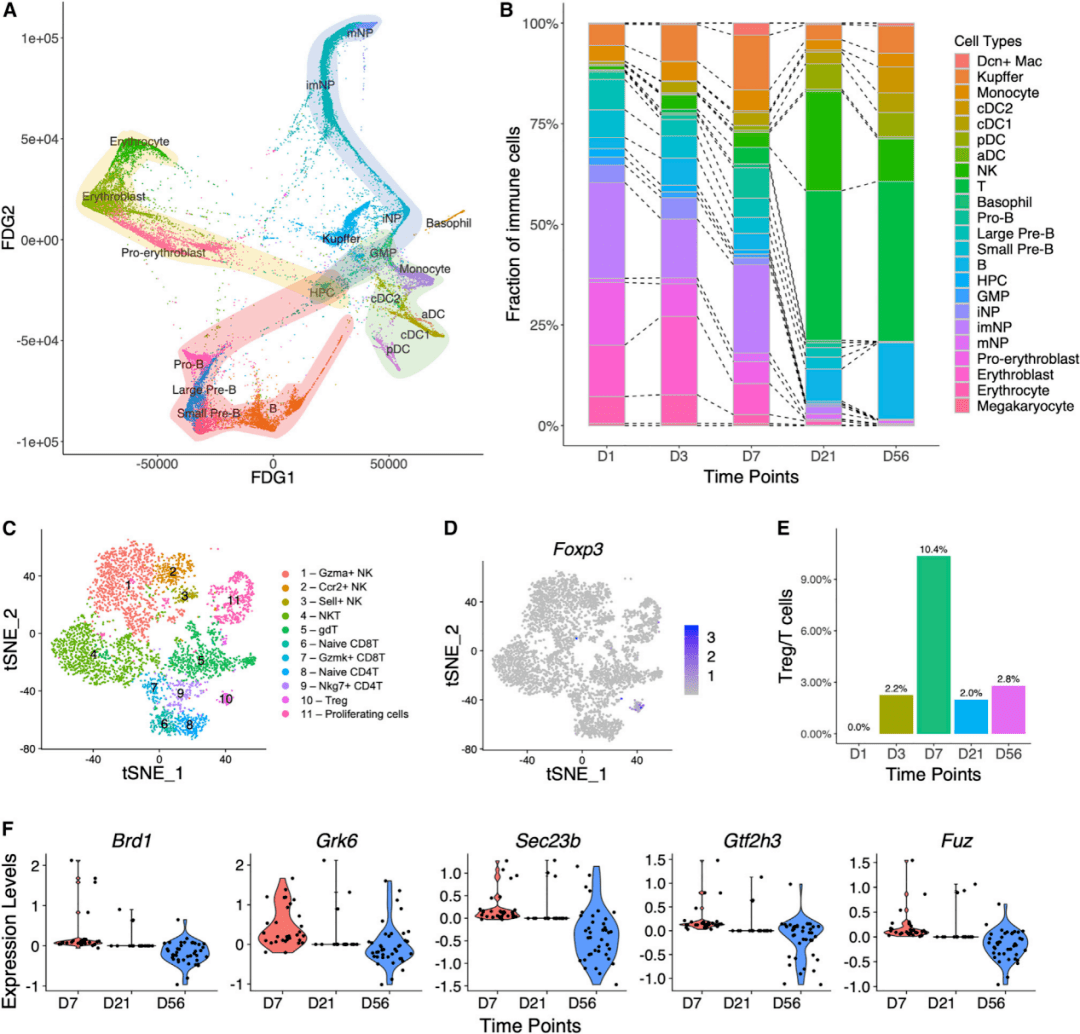
图5 造血和免疫细胞群的动态变化
在UMAP上,一个Kupffer细胞簇位于树突状细胞(dendritic cells,DCs)和单核细胞附近(图1A)。该簇表达Kupffer细胞标记物Adgre1和Csf1r(图1B、6A、6B),在D7达到峰值(图6D)。在单独分析Kupffer细胞时,研究人员鉴定出6种亚型KC1~KC6,它们具有不同的基因表达谱和TF活性(图6C)。此外,研究人员识别出一个巨噬细胞亚群(Dcn+ Mac),其特征是高表达核心蛋白聚糖,这是富含亮氨酸的小蛋白多糖家族的成员。除了传统的Kupffer细胞标记物Adgre1和Csf1r(图1B、6A、6B和6E)外,Dcn+ Mac还表达一般的EC标记物Pecam1、Eng、Kdr,以及LSEC标记物Lyve1和Clec4g,尽管MaVEC标记物Vwf为阴性(图1B、6E和6F)。几乎所有的Dcn+ Mac都出现在D7(图6D),这促使研究人员进一步对这个独特的亚型进行深入研究。
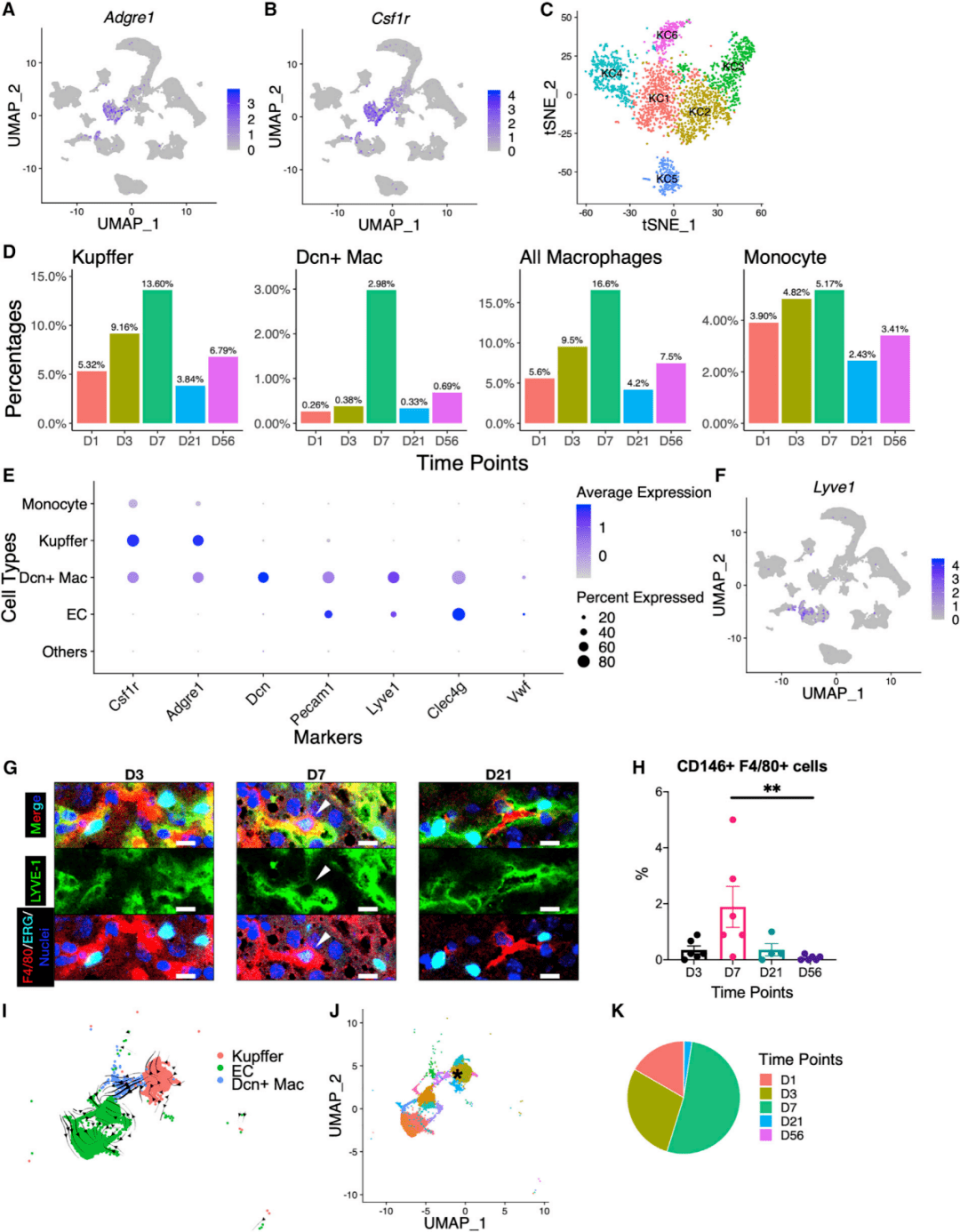
图6 巨噬细胞的一个亚型在出生后第7天短暂出现
通过LSEC标记物LYVE-1、内皮细胞特异性TF、ERG和巨噬细胞标记物F4/80的免疫共染,验证了Dcn+ Mac的存在及其在D7的丰度(图6G)。FACS分析也检测到这种细胞类型,并在D7验证了其峰值水平(图6H)。RNA速度分析显示,Dcn+ Mac的发育轨迹来自Kupffer细胞而非ECs(图6I)。研究人员通过噬菌体画像绘制了Dcn+ Mac中显著上调的前10个基因。过渡点的Kupffer细胞群(过渡KC)在D1、D3和D7被鉴定(图6K),其在新生期显示出分化潜能。在过渡KC和其他远离Dcn+Mac的Kupffer细胞之间进行差异基因表达分析,研究人员鉴定了在过渡KC中选择性表达的基因,如Fcna、Marco、Cd163、Cd209f等(图6J)。通过CellPhoneDB的配体-受体分析,研究人员确定了与Dcn+ Mac相互作用的细胞类型,包括HSCs、成纤维细胞、间皮细胞、胆管上皮细胞、VSMCs和所有ECs亚型。这些细胞类型与Kupffer细胞之间的相互作用较少(图7A)。然后,研究人员检查了与Dcn+ Mac相关的信号串扰,并与Kupffer细胞进行了比较(图7B),揭示了Treg在D7积累的意义,以及Treg与Dcn+ Mac细胞之间潜在的串扰。
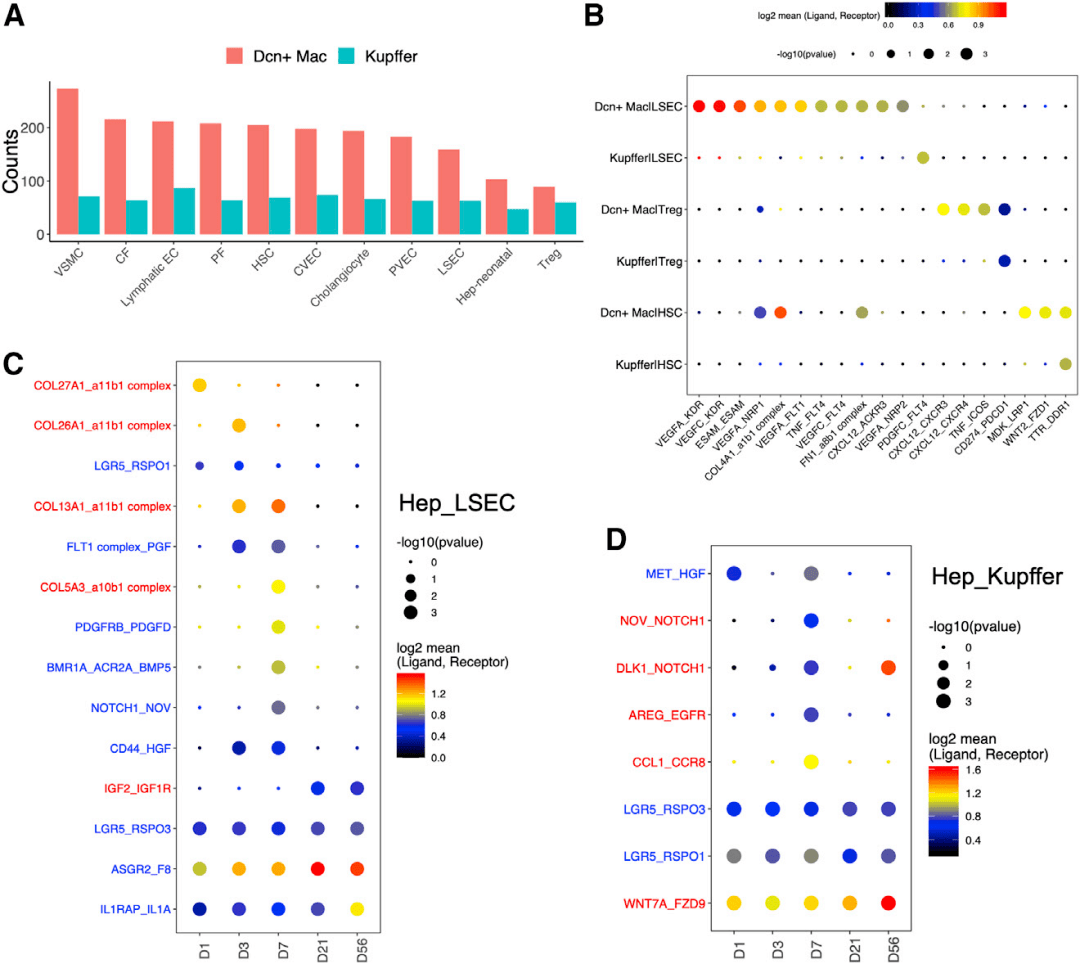
图7 预测肝细胞-细胞相互作用
6. 肝细胞和NPC之间的细胞间通讯
使用上述相同的方法,研究人员分析了肝细胞和NPCs之间的串扰,以及出生后早期细胞间信号传导的动态变化。分别对所有5个时间点进行配体-受体分析,鉴定了LSECs和肝细胞之间的RSPO1/3-LGR5相互作用(图7C)。RSPO3-LGR5在所有时间点均显著表达,而RSPO1-LGR5仅在新生儿期表达。RSPO与LGR5的结合,存在于近中心的肝细胞上,促进Wnt信号传导,调节近中心的区的分区代谢活动。在所有时间点的Kupffer细胞和肝细胞之间也发现了这一信号事件(图7D)。研究人员发现,对于几乎所有与肝细胞密切相互作用的细胞类型,与其他时间点相比,最活跃的串扰发生在D7。一些相互作用对,包括肝细胞与LSECs之间的PDGFRB-PDGFD(图7C)、肝细胞与Kupffer细胞之间的NOV-NOTCH1 (图7D),也仅在D7被鉴定。综上,出生后肝脏中的一个关键时间点D7,涉及许多分子和细胞活动和通讯。
结论
本研究在单细胞分辨率水平上详细地描述了小鼠新生儿肝脏如何发育成一个主要的代谢器官。轨迹分析显示,在代谢区域构建中肝细胞和LSECs的发育、协同发育和功能成熟。肝细胞类型之间复杂的相互作用显然参与了早期阶段的协调发育程序。此外,研究人员在D7鉴定了一组特殊的巨噬细胞,表现出巨噬细胞和内皮细胞的混合表型。细胞间相互作用分析表明,Dcn+ Mac可能在调节窦状血管化和Treg细胞中发挥作用。
转自:华大时空
- 本文固定链接: https://maimengkong.com/kyjc/1496.html
- 转载请注明: : 萌小白 2023年4月29日 于 卖萌控的博客 发表
- 百度已收录
