基因表达的改变是植物适应环境变化的一种机制。热是一种非生物胁迫,可以在许多方面影响植物。平均温度的微小变化会导致平均产量的降低。繁殖期间严重的热胁迫事件会导致结实率的重大变化。营养早期的热胁迫事件会导致叶片衰老和生长速率降低。植物转录组分析发现,许多基因在非生物胁迫下表现出转录物丰度的变化。热休克因子(HSF)转录因子(tf)在热胁迫下被激活,并通过与顺式作用元件的相互作用调节许多基因的表达水平。先前的研究已经发现,在拟南芥、玉米和水稻中,HSF结合位点基序在上调基因上富集。一些HSF稳定表达,并在热胁迫下被激活。其他一些转录因子和顺式调控元件也参与了对热胁迫的反应。先前对热胁迫引起基因表达改变的潜在机制的了解,使这个系统成为研究热胁迫变量反应的一个很好的系统。此外,这为考虑特定分子反应的基因型x环境(GxE)相互作用提供了一个有用的系统。GxE的相互作用对于在波动环境中开发高生产力的作物品种非常重要,但我们对特定环境中驱动GxE的分子机制的了解往往有限。阐明基因表达调控机制的一个困难是我们识别和注释影响基因表达模式和水平的顺式调控元素的能力。在某些情况下,研究人员简单地利用转录起始位点上游的所有区域(通常为1-2kb)来寻找潜在的基序或调控元件。近年来,许多研究小组专注于利用染色质特性来识别可能对调控很重要的保守非编码序列(CNSs)。具体来说,可接近性染色质区域或特定的组蛋白修饰可用于记录功能重要的顺式调控元件。然而,对于对非生物胁迫的基因表达反应,尚不清楚有多少比例的顺式调控元件被预先标记(在胁迫前可接近)或仅在胁迫暴露后表现出可及性的改变。
野生物种的局部适应和提高作物植物恢复能力的育种可能利用基因表达对非生物胁迫反应的自然变异。这种变异可以归因于转录因子等反式作用因子的不同活性,或由于顺式作用调节变异可以改变特定基因的调节方式。我们对可变顺式调节元件的进化来源的理解是有限的。基因获得或失去对环境刺激反应的机制尚不清楚。假设大部分顺式调控信息是通过转录因子结合位点(TFBSs)提供的。虽然单核苷酸多态性(SNPs)可以改变TFBS序列,导致顺式调控信息的改变,但尚不清楚新的TFBS和对特定环境的响应的通路是如何产生的。插入/删除(InDel)多态性,包括转座元件(TE)插入,也可能在改变顺式调控元件的组成方面起着重要作用。一些研究比较了不同基因型对热胁迫的基因表达反应。虽然许多基因在不同基因型中对热胁迫表现出相似的反应,但也有一些基因表现出显著的等位基因变异的反应。利用杂交基因型和等位基因特异性表达分析发现了顺式和反式调控变异的证据。等位基因的比较表明,结构变异可能在驱动变量响应性方面发挥重要作用,但这尚未得到详细的研究。

近日,美国明尼苏达大学在著名期刊《 Genome Biology》发表了题为“ Mapping responsive genomic elements to heat stress in a maize diversity panel”的研究论文。在本研究中,研究者利用102个在对照和热胁迫条件下生长的玉米基因型的重测序,研究了基因表达的变化和与热胁迫响应相关的基因组调控元件,
利用基因组和转录组数据来记录eQTLs及其调控基因,同时发现了影响对热胁迫的可变响应性的顺式eQTL,以记录热诱导表达的等位基因变异。这种方法被证明是一种有效的方法来记录数以百计的顺式等位基因变异的热胁迫反应。许多热响应基因在热胁迫条件下可及性染色质和转录因子占用率发生了变化。这两种方法的结合为识别高质量的候选因果区域提供了机会,可以解释热响应性的等位基因变异。
与光合作用参数相关的全基因组标记
102个不同玉米基因型之一的组,包括刚性秸秆、非刚性秸秆和iodent亚群,此组代表了玉米威斯康星州多样性 (WiDiv),被选择用于表征玉米对热胁迫的基因型响应(图1a)。该种群使用了1,132,322个变异集(1,032,834个snp + 99,488个InDels)。研究者在这项研究中使用的基因型包括3到32个自交系,每个自交系来自7个玉米亚群体(图1a)。用于从较大的WiDiv亚群中选择自交系的子集的选择标准保留了高水平的多样性,同时减少了总体的种群结构。根据这些已知的子种群,前两个主要成分(PCs)确实提供了一些分离。从基因型矩阵中计算出的前5个PCs解释了所选102个基因型中17.1%的群体结构变异,并在后续分析中作为协变量控制群体结构。
将这些基因型的幼苗(播种后14天)置于40°C的4小时热胁迫下。每株植物的第三片叶子还评估了三个基于叶绿素荧光的参数,包括:(1)PSII的光适应有效量子产率[Y(II)]和两条非光化学耗散路径,(2)PSII的非调节能量耗散[Y(NO)],以及(3)对照和热胁迫条件下PSII的调节能量耗散[Y(NPQ)]。较高的Y(II)值表示光合胁迫较小,较高的Y(NPQ)值表示持续的光化学胁迫通过类胡萝卜素能量耗散得到缓解,较高的Y(NO)值表示持续的光化学胁迫没有得到缓解。平均而言,与对照条件相比,在热胁迫条件下,植物表现出较低的Y(NO)值和较高的Y(II)值,而记录的Y(NPQ)中心在对照和热胁迫条件之间没有显示出显著的变化(图1b)。3个性状[Y(II)、Y(NPQ)和Y(NO)]在对照中均表现出较高的广义遗传力(H2)(0.692、0.682和0.716),高于热胁迫植物(0.538、0.563和0.642),表明热胁迫条件下这些性状的变异幅度高于对照植物。
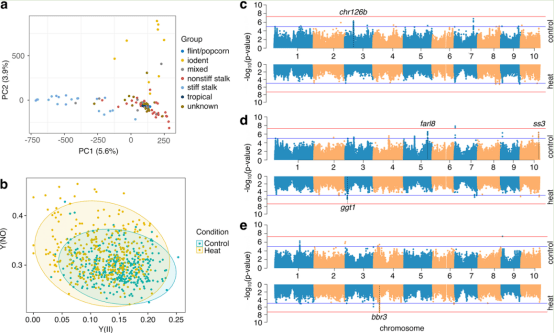
图1玉米多样性组对热胁迫的表型响应
WiDiv组的一个子组对热胁迫的转录组响应
研究者继续评估在同一组基因型中对热胁迫事件的转录反应。幼苗(播种后14天)置于对照条件或4小时的热胁迫(40°C),并从每个处理/基因型组合的2-3个个体的池中收集叶片组织用于RNA-seq。RNA-seq数据与玉米B73 AGPv4参考基因组进行比对,所有样本均表现出较高的总体比对率,从84.5到98.6%。在删除了所有样本中没有读取数的基因后,得到了一个基因计数矩阵,其中包含分配给39,511个玉米AGPv4基因模型的读取数。为了评估采集过程中样品之间的一致性,在每种条件的早期、中期和后期采集时间点分别采集B73的3个生物重复。对照组和热胁迫组重复之间的平均两两Spearman相关性分别为0.987和0.972,表明采集过程中样品具有较高的重复性。使用主成分分析评估所有样本的转录组的整体变异性(图2a)。基于处理,第一主成分分离样品,表明热胁迫的显著影响(图2a)。第二个主成分提供了基于基因型的适度分离,但没有表明已知种群结构的主要贡献(图2a)。假设,对热胁迫反应的转录组移位较少(基于主成分1)的基因型可能在对照和热条件下表现出更相似的光合参数BLUPs分布。PC1在对照和加热条件下的绝对差异反映了每个基因型的转录组可塑性,可用于将基因型分为大、中、小转录组响应。每个基因型的BLUP被确定为基因型效应后,控制实验因素的测量表型从任何条件。研究者计算了对热胁迫有大、中、小转录组反应的基因型在对照和热条件下的BLUPs之间的相关性(图2b)。总的来说,对于每一个测量的表型,观察到相关系数的增加与更大的转录组可塑性,这表明表型和转录组反应之间的正相关。对B73中对热响应显著上调的基因的GO(基因本体论)富集分析显示,几个显著的GO富集与热响应功能相关,为B73样本中强烈的热胁迫反应提供了证据。
对整个数据集的分析显示,在大多数基因型中,2628个基因对热的响应表现出一致的上调。对这2628个基因对热胁迫的平均响应和热胁迫表达水平的变异系数(CV)的比较突出了对不同基因型的响应具有强烈表达响应和有限变异性的基因子集(图2c)。许多在基因型间具有最高表达响应和相对低变异的基因先前被鉴定为热休克蛋白(HSPs)。在maizeGDB数据库(https://www。maizegdb.org),其中25个HSPs在热胁迫下一致上调,表达值高,cv低(图2c)。另外6个未被注释为HSPs的基因(Zm00001d004243、Zm00001d022630、Zm00001d031436、Zm00001d033990、Zm00001d039933和Zm00001d048592)对热胁迫也表现出强烈和一致的反应,可能在热胁迫反应中发挥重要作用。热休克因子(HSFs)是一类已知的转录因子,在热胁迫和激活热休克因子的基因表达响应中起着重要作用。在之前报道的31个玉米非冗余HSFs中,有17个在数据集中显示了可检测到的表达。其中8个HSF (ZmHsf02, ZmHsf25, ZmHsf05, ZmHsf08, ZmHsf26, ZmHsf11, ZmHsf03和ZmHsf13)在100个基因型中对热的响应一致上调(图2d)。有趣的是,在本研究中使用的所有基因型中,ZmHsf06都被下调,并且在之前的研究中B73在多个时间点对热胁迫的响应中表现出表达减少。结果表明,热胁迫事件在这一多样化的群体中导致了对热胁迫的稳健响应,并为进一步研究更多不同基因型的可变响应提供了机会。
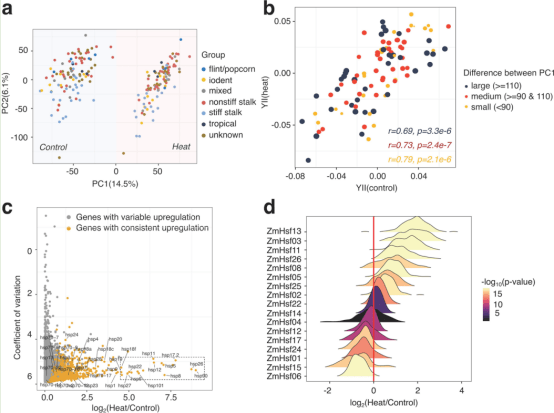
图2研究玉米基因型在高温和对照条件下的转录组多样性
热胁迫下可变基因表达响应的表征
为了在该图谱中绘制可能影响基因表达水平或热胁迫反应的调控变异,我们在至少10%的基因型中保留了每百万分数(CPM) > 1的基因,结果分别有20,255和20,306个基因在对照和热条件下表达。在这些基因中,19,642个基因在两种条件下共同表达,而613和664个基因在对照或热条件下唯一表达。在热条件下唯一检测到的664个基因中,有3个是热休克因子(ZmHsf03, ZmHsf11和ZmHsf26)。我们分别使用对照或热表达数据进行eQTL定位,以绘制影响表达基因转录水平的基因组元素。位于距离目标基因1Mb内的显著相关snp被归为顺式eQTL,而位于距离目标基因1Mb的>的snp被归为反式eQTL。顺式eQTL的1Mb截断位点已经在玉米eQTL的研究中使用过,该截断位点在我们的群体中具有非常低的连锁不平衡(r2<0.05)。通过计算基因组中每个10 kb窗口中显著的反式相互作用的数量来评估潜在的有效反式eQTL。在热胁迫和对照条件下,通常检测到几个至少有10个互作的染色体区域。在9号染色体上检测到3个10 kb的区域在热胁迫下与基因表达有远距离的相互作用。然而,检测到的“热点”的靶点数量有限(<20),本研究中检测跨eQTL热点的能力可能受到样本量的限制。与之前的研究一致,本研究中发现的顺式eQTL在对照和热条件下比反式eQTL能解释更多的基因表达方差。顺式eQTL分析分别在对照和热胁迫条件下鉴定出10548和10391个顺式eQTL调控基因(eGenes)。在仅在热胁迫条件下表达的基因中检测到的顺式eQTL特别有趣,因为它们反映了热胁迫下基因表达激活的可变水平。在664个仅热表达(heo)基因中,有518个基因具有显著的顺式eQTL (heo-cis-eQTL),我们将这些基因称为heo-egenes。Heo-eGenes包括许多转录因子,如zhd20、iaa37、abi33、gata17、sbp18、wrky35、bhlh110、dof27和ereb196。此外,两个热休克因子ZmHsf03和zmhsf26被检测为heo-eGenes。
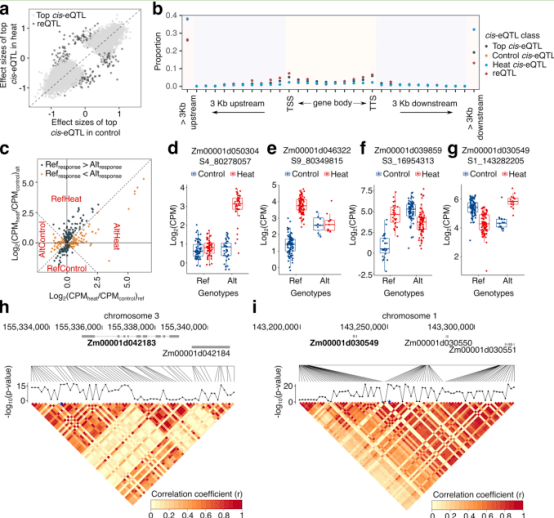
在玉米多样性图谱中检测到响应基因和eQTLs
本研究同时也检测与玉米热响应相关的染色质结构变化以及瞬时表达测定证实了热反应的等位基因变异,在此不多一一赘述。
植物经常遇到非生物胁迫,有证据表明许多物种对胁迫的反应是可变的。本研究着重于描述热胁迫事件的基因表达反应的自然变异来源。研究重点是在40°C下进行中等(非致命性)4小时热胁迫处理。虽然许多基因对热胁迫反应迅速(<30分钟),但在早期时间点的反应有相当动态的变化。为了尽量减少因采样时间超过15分钟而引起的潜在影响,研究者重点进行了中间4小时的热胁迫处理。该研究调查的群体中,有许多基因对热胁迫有高度保守的反应。许多对热胁迫事件有强烈、一致反应的基因是热胁迫蛋白质,它们可能起到伴侣的作用。这证实了本研究中使用的基因型面板都能够产生强烈的热响应。虽然基因型也可以对热胁迫作出反应,但有许多基因具有不同的反应,这为在特定非生物胁迫背景下通过环境(GxE)相互作用研究基因型提供了机会。
- 本文固定链接: https://maimengkong.com/zixun/1507.html
- 转载请注明: : 萌小白 2023年5月2日 于 卖萌控的博客 发表
- 百度已收录
